
Plant growth regulators (PGR s) are molecules that influence the development of plants and are generally active at very low concentrations. There are natural regulators, which are produced by the plant itself, and also synthetic regulators; those found naturally in plants are called phytohormones or plant hormones.
Substances considered phytohormones include auxins, gibberelins, cytokinins, abscisic acid and ethylene, and more recently brassinosteroids, salicylic acid, jasmonates, systemin, polyamines, nitric oxide and signal peptides.
There are differences between plant and animal hormones. For example, animal hormones are synthesised in particular organs or tissues, and by definition they act in different places to where they are produced. This is not necessarily true for phytohormones; some exert their action in exactly the same place where they are synthesised.
Although all phytohormones have their own specific effects, their combination produces a varied response in plants.

Auxins
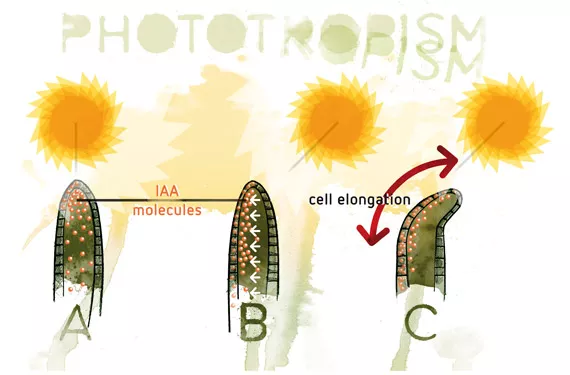
The main effect of auxins is to cause cell elongation, mainly due to the alteration of cell wall plasticity. Auxins are synthesised in the apical meristems and to a lesser degree in the roots. The main auxin to be synthesised naturally by plants is indole acetic acid (IAA), although others have been found such as phenylacetic acid, the chlorindoles and, more recently, indole butyric acid (IBA). The movement of these phytohormones is from the apices to the roots (basipetal) and vice versa (acropetal). However, basipetal movement is much more rapid than acropetal movement.
Some of the effects of auxins in plants include:
Apical dominance. It is well known among growers that when one eliminates the main apical axis (main vertical stem) of a plant, secondary apices will begin to grow and several of these will go on to form main stems. This occurs because the auxins produced by the apical meristem suppresses the growth and development of secondary buds.
Rhizogenesis. Auxins are the main components responsible for the formation of root cells. This property is used by gardeners to produce cuttings: applying auxins to the base of the cut promotes the formation of new roots. This rhizogenesis occurs at very low concentrations of auxins, since higher concentrations of auxins suppress root growth and development. However, it is the presence of other phytohormones that determines whether the new cells become roots or other organs. The balance between auxins and cytokinins plays a very important role in this process. Thus when plant cells are grown in vitro in culture media, if the concentration of auxins is greater than that of cytokinins, new roots will be formed. However, if the concentration of cytokinins is greater than that of auxins, the cells will eventually develop into new buds. When the concentration of the two hormone types is similar, cell growth will occur without differentiation, forming a mass of developing cells called a callus.
Geotropism. Gravity exerts an effect on plant development. When a plant stem is placed in a horizontal position, lateral buds will begin to develop and may form roots in the zone which is in contact with the soil. This is due to the accumulation of auxins due to the effect of gravity. This phenomenon is used to obtain new plants using a technique called layering.
Phototropism. Plants tend to grow towards the light. This process is regulated by auxins, which accumulate in parts that receive less light; this results in the elongation of the cells in this zone and makes the stem curve towards the light.
Regulation of abscission. Abscission is the shedding of some parts of the plant. In many cases the cause is the ageing of the plant tissue, called senescence. The exogenous application of auxins will reduce abscission in many species.
Fruit set. When pollination and fecundation occur, the concentration of auxins in the fruit usually increases, possibly as a result of production by the developing seeds. If fecundation does not occur, the fruit are shed instead of developing and maturing. But by applying auxins, the formation and maturation of fruit can occur without pollination or fecundation (and therefore seed formation) being necessary. The development of fruit without fecundation is called parthenocarpy and it is widely used when the formation of seeds is undesirable or when no pollination is possible. This occurs when insect-pollinated plants are grown into greenhouses. When there are no pollinating insects, exogenous auxins are applied to promote fruit set.
Gibberelins
These phytohormones are partly responsible for cell division and the elongation of stems and other tissues. They were discovered by Japanese researchers studying a disease in rice. The disease caused recently germinated seedlings to acquire a yellowish colour and the stem to elongate excessively, leading to the death of the plant. The researchers discovered that these symptoms were caused by the Gibberella fujikuroi fungus. This fungus produces a large quantity of these phytohormones which are introduced into the host plant.
Since then, various types of gibberelins have been discovered and isolated. These were given successive numbers as they were discovered; GA1, GA2, GA3, etc. GA3 is gibberellic acid.
Gibberelins are synthesised mainly in meristematic organs and developing tissues.
Functions of gibberelins
Seed germination. In seeds, some of the gibberelins combine with glucosides, and become inactive in this form. During germination, enzymes destroy this combination and the gibberelins are unlocked and activated. This stimulation of germination has been demonstrated in a number of experiments which showed how the application of gibberelins accelerated the germination of lettuce seeds. It was also shown that exposure to light accelerated the germination of lettuce seeds. Later studies showed that light accelerates the transformation of the gibberelins from the inactive conjugated form to the active form.
Sex expression. In species with unisexual flowers - that is, separate male and female flowers, either on the same plant (monoecious) or on different individuals (dioecious) - gibbberelins appear to have a regulatory effect on sex expression. For example, the application of gibberelins in female asparagus plants produces the appearance of male and hermaphrodite flowers. By contrast, gibberellin application in maize plants produces the appearance of female flowers in the tassels (masculine inflorescences).

Influence during the juvenile period. Juvenile plants are different to adult plants - for example, developing fruit trees must mature for several years after seed germination before they are capable of producing flowers and fruits. In some cases they also have different characteristics when adult (for example, the presence of spines or leaves with different shapes). Gibberelins play an important role in the transition from the juvenile period to the adult period. In some plants, such as ivy, the exogenous application of gibberelins produces the expression of branches with juvenile characteristics.
Fruit set. As with auxins, gibberelins stimulate fruit set in some species.
Induction of flowering. Some plants require long days or cold periods to flower, but the application of gibberelins induces flowering independently of the photoperiod or the temperature.
Cytokinins
The discovery of these phytohormones was due mainly to in vitro cultivation studies. The first observation was that “coconut milk” (the endosperm of the fruit) promoted the growth of several tissues cultivated in vitro.
The first natural cytokinin isolated and identified was named zeatin, since it was isolated from maize (Zea mays) seeds.
The main function of the cytokinins is to produce cell division and retard senescence. As mentioned above, cytokinins in combination with auxins lead to the formation of undifferentiated cell masses called calluses. They also stimulate the development of lateral apices when applied exogenously, breaking apical dominance.
Ethylene
Ethylene is a simple hydrocarbon that is a gas under normal conditions. The effects of ethylene on plants was discovered when streets were illuminated with carbide lamps. The combustion process led to the emission of ethylene and the leaves of trees near these lamps turned yellow and were shed.
The main function of ethylene is on fruit maturation and the senescence of leaves and flowers. In species with climacteric fruit, maturation is induced by an increase in this hormone. It is also responsible for the change in colour of some non-climacteric fruits (that is, fruit whose maturation is not affected by ethylene), as occurs in citrus fruit. Ethylene is used to mature fruit that has been collected prematurely. It is applied by burning in closed chambers or with ethephon, a product which decomposes into ethylene when hydrolysed in the plant.
Another function attributed to both ethylene and gibberelins is the regulation of sexual expression in dioecious plants. The application of ethylene in asparagus induces the appearance of female flowers in male plants.
Ethylene plays an important role along with jasmonic acid in stimulating the production of substances that protect the plant from biotic and abiotic stresses.
Abscisic acid (ABA)
As the name indicates, this hormone is directly implicated in the senescence and abscission of leaves, flowers and fruits. It also affects the latency of some seeds.
As in the case of ethylene, this phytohormone induces the expression of resistance genes for a number of stresses. One effect of ABA is to produce stomatal closure during drought conditions, thus preventing dehydration in the plant.

